
Expression analysis of G Protein-Coupled Receptors in mouse macrophages
- Research
- Open Access
Expression analysis of G Protein-Coupled Receptors in mouse macrophages
- Received: 29 January 2008
- Accepted: 29 April 2008
- Published: 29 April 2008
Abstract
Background
Monocytes and macrophages express an extensive repertoire of G Protein-Coupled Receptors (GPCRs) that regulate inflammation and immunity. In this study we performed a systematic micro-array analysis of GPCR expression in primary mouse macrophages to identify family members that are either enriched in macrophages compared to a panel of other cell types, or are regulated by an inflammatory stimulus, the bacterial product lipopolysaccharide (LPS).
Results
Several members of the P2RY family had striking expression patterns in macrophages; P2ry6 mRNA was essentially expressed in a macrophage-specific fashion, whilst P2ry1 and P2ry5 mRNA levels were strongly down-regulated by LPS. Expression of several other GPCRs was either restricted to macrophages (e.g. Gpr84) or to both macrophages and neural tissues (e.g. P2ry12, Gpr85). The GPCR repertoire expressed by bone marrow-derived macrophages and thioglycollate-elicited peritoneal macrophages had some commonality, but there were also several GPCRs preferentially expressed by either cell population.
Conclusion
The constitutive or regulated expression in macrophages of several GPCRs identified in this study has not previously been described. Future studies on such GPCRs and their agonists are likely to provide important insights into macrophage biology, as well as novel inflammatory pathways that could be future targets for drug discovery.
Keywords
- RAW264 Cell
- Macrophage Population
- Unstimulated Macrophage
- Bone Marrow Hematopoietic Stem Cell
- Macrophage Biology
Background
Macrophages are key cellular mediators of acute and chronic inflammation. They can be defined on the basis of morphology, function (e.g. non-specific uptake of particles) and expression of specific cell surface markers (e.g. EMR1 that is detected by the F4/80 antibody). EMR1 belongs to the GPCR super-family [1], and it is clear that macrophages express a diverse repertoire of other GPCRs [2]. A large number of GPCRs are yet to be deorphanized and hence, there is great potential for this family in both the discovery of gene function and in drug development. In this study we performed a systematic micro-array analysis of GPCR expression in primary mouse macrophages and identified family members that are either enriched in macrophages compared to a panel of other cell types, or are regulated by the pro-inflammatory stimulus, bacterial LPS. Some of these GPCRs are likely to provide future targets for drug discovery in inflammatory disease settings.
Results
Constitutive GPCR expression in mouse macrophages
Micro-array expression profiling across a panel of mouse and human tissues has enabled detailed analysis of tissue-specific gene expression [3]. In this study, we analyzed the constitutive and regulated expression of GPCRs in macrophages by interrogating a more detailed micro-array dataset of 91 murine cell types and tissues [3, 4]. This data, available through NCBIs Gene Expression Omnibus ([5] accession number GSE10246), can also be used to probe gene expression profiles for an extensive array of other cell types.
The cell and tissue panel included two of the most commonly used primary mouse macrophage models; bone marrow-derived macrophages (BMM) and thioglycollate-elicited peritoneal macrophages (TEPM), as well as microglia (a resident tissue macrophage population of the brain) and the widely utilized macrophage-like cell line, RAW264. We analyzed the expression patterns of all GPCRs as defined by the International Union of Basic and Clinical Pharmacology [6, 7]. We then compared the GPCR expression profiles of BMM and TEPM to the remaining panel of cell types, and in doing so, identified 67 GPCRs that were detectably expressed either constitutively or in an LPS-induced manner in these primary macrophage populations [see additional file 1].
Summary of GPCRs expressed by macrophages.
Highly Expressed | |
|---|---|
BMM |
TEPM |
|
22 10 only enriched in BMM |
23 11 only enriched in TEPM |
12 regulated in both BMM and TEPM | |
LPS Regulation | |
|---|---|
Induced | |
BMM |
TEPM |
|
27 14 only regulated in BMM |
16 3 only regulated in TEPM |
13 regulated in both BMM and TEPM | |
Repressed | |
|---|---|
BMM |
TEPM |
|
19 9 regulated only in BMM |
18 8 regulated only in TEPM |
10 regulated in both BMM and TEPM | |
Macrophage-enriched GPCRs.
Relative Normalised Expression |
|||||
|---|---|---|---|---|---|
Receptor Class |
Accession Number |
Gene Name |
BMM |
TEPM |
RAW264.7 |
Enriched in both BMM and TEPM | |||||
Class A |
|||||
NM_009779 |
C3ar1 |
1361.1 |
683.6 |
58.3 |
|
NM_007577 |
C5r1 |
822.6 |
25.6 |
10.5 |
|
NM_183168 |
P2ry6 |
434.0 |
94.0 |
196.4 |
|
NM_009917 |
Ccr5 |
214.2 |
20.5 |
0.6 |
|
NM_021476 |
Cysltr1 |
208.9 |
12.5 |
40.6 |
|
NM_004230 |
Edg5 |
30.2 |
69.3 |
6.6 |
|
NM_008967 |
Ptgir |
24.7 |
72.1 |
1.7 |
|
NM_009924 |
Cnr2 |
14.2 |
27.2 |
6.7 |
|
NM_009912 |
Ccr1 |
14.1 |
235.1 |
16.3 |
|
NM_145700 |
Ccrl1 |
11.1 |
26.6 |
12.7 |
|
NM_008964 |
Ptger2 |
10.7 |
34.4 |
0.8 |
|
Class B |
|||||
NM_010130 |
Emr1 |
190 |
22.2 |
28.3 |
|
Enriched in BMM | |||||
|---|---|---|---|---|---|
Class A |
|||||
NM_183031 |
Ebi2 |
113.5 |
6.1 |
245.2 |
|
NM_009915 |
Ccr2 |
92.1 |
1.2 |
1.3 |
|
NM_030720 |
Gpr84 |
77.5 |
0.6 |
2.7 |
|
NM_008042 |
Fprl1 |
32.0 |
1.0 |
0.8 |
|
NM_008152 |
Gpr65 |
29.3 |
9 |
6.6 |
|
NM_009910 |
Cxcr3 |
26.3 |
1.1 |
7.6 |
|
NM_007420 |
Adrb2 |
22.1 |
1.3 |
9.3 |
|
NM_145066 |
Gpr85 |
13.7 |
1.7 |
7.6 |
|
NM_009987 |
Cx3cr1 |
11.3 |
0.7 |
61.4 |
|
Class B |
|||||
NM_018782 |
Calcrl |
45.6 |
1.1 |
6.1 |
|
Enriched in TEPM | |||||
|---|---|---|---|---|---|
Class A |
NM_009911 |
Cxcr4 |
4.3 |
178.9 |
0.6 |
NM_017466 |
Ccrl2 |
3.1 |
154.9 |
1.1 |
|
NM_021381 |
Prokr1 |
1.0 |
36.6 |
0.7 |
|
NM_022320 |
Gpr35 |
2.5 |
22.0 |
0.4 |
|
NM_008311 |
Htr2b |
1.1 |
19.9 |
0.5 |
|
NM_008773 |
P2ry2 |
8.5 |
19.2 |
1.5 |
|
NM_133200 |
P2ry14 |
4.0 |
18.4 |
0.4 |
|
NM_175493 |
Gpr68 |
0.9 |
17.9 |
9.3 |
|
NM_008772 |
P2ry1 |
4.4 |
10.6 |
0.6 |
|
Class B |
|||||
NM_011925 |
Cd97 |
0.4 |
22.9 |
0.2 |
|
Class C |
|||||
NM_022420 |
Gprc5b |
0.6 |
60.6 |
0.6 |
|
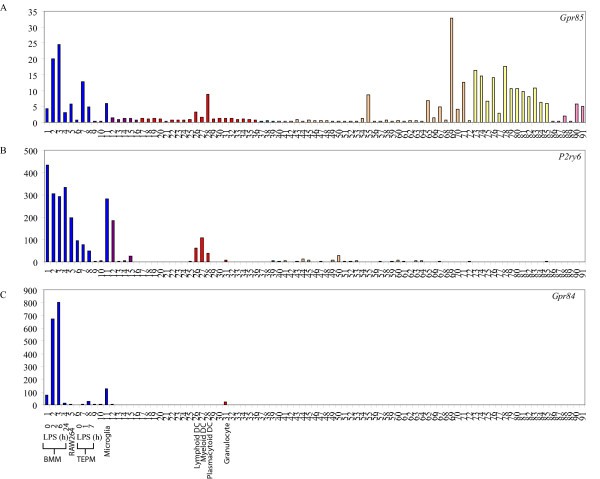
Macrophage-restricted GPCRs. Micro-array analysis of Gpr85 (A), P2ry6 (B) and Gpr84 (C) mRNA expression across a panel of 91 murine cell types and tissues. Data points show normalised values and similar cell types are grouped according to bar colour; blue indicates primary macrophage cell types, purple indicates bone-related cell types, red indicates other immune cell types, green indicates stem cell populations, orange indicates whole tissue samples, yellow indicates neuronal and retinal cell types and pink indicates cell lines. Additional file 3 gives details of the 91 cell types and tissues profiled.
Differential GPCR expression in distinct macrophage populations
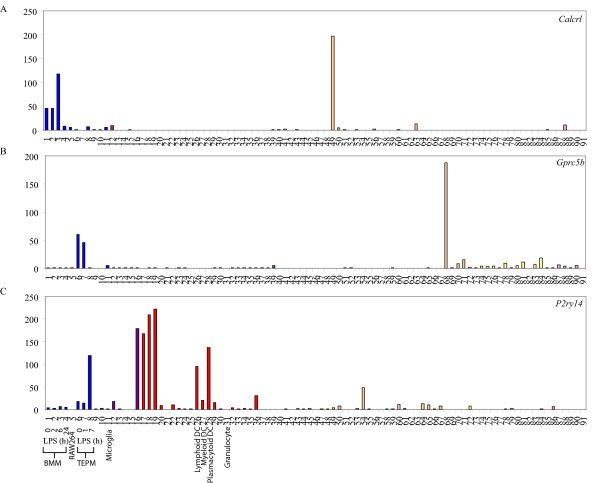
GPCRs differentially regulated in BMM and TEPM. Micro-array analysis of Calcrl (A), Gprc5b (B) and P2ry14 (C) mRNA expression across a panel of 91 murine cell types and tissues. Data points show normalised values and similar cell types are grouped according to bar colour; blue indicates primary macrophage cell types, purple indicates bone-related cell types, red indicates other immune cell types, green indicates stem cell populations, orange indicates whole tissue samples, yellow indicates neuronal and retinal cell types and pink indicates cell lines. Additional file 3 gives details of the 91 cell types and tissues profiled.
LPS-regulated GPCR expression in mouse macrophages
GPCRs induced by LPS in macrophages.
Relative Normalised Expression |
|||||||
|---|---|---|---|---|---|---|---|
Receptor Class |
Accession Number |
Gene Name |
BMM |
TEPM |
|||
2 h |
6 h |
24 h |
1 h |
7 h |
|||
Induced by LPS in BMM and TEPM | |||||||
Class A |
|||||||
NM_009630 |
Adora2a |
1.2 |
28.2 |
3.3 |
4.2 |
52.2 |
|
NM_007413 |
Adora2b |
27.4 |
190.9 |
17.4 |
3.0 |
3.3 |
|
NM_007719 |
Ccr7 |
1.0 |
5.1 |
1.9 |
0.4 |
2.9 |
|
NM_017466 |
Ccrl2 |
39.2 |
65.7 |
46.7 |
13.8 |
9.2 |
|
NM_010336 |
Edg2 |
7.3 |
5.7 |
0.3 |
1.0 |
10.0 |
|
NM_007904 |
Ednrb |
6.7 |
2.0 |
10.3 |
0.7 |
5.3 |
|
NM_013521 |
Fpr1 |
5.9 |
120.9 |
23.3 |
1.0 |
21.4 |
|
NM_008042 |
Fprl1 |
3.8 |
41.6 |
28.6 |
3.1 |
570.6 |
|
NM_030701 |
Gpr109b |
13.6 |
48.2 |
5.6 |
10.8 |
212.0 |
|
NM_030720 |
Gpr84 |
8.7 |
10.3 |
0.2 |
7.8 |
47.5 |
|
NM_145066 |
Gpr85 |
3.6 |
5.6 |
0.5 |
18.1 |
4.9 |
|
Class B |
|||||||
NM_018782 |
Calcrl |
1.0 |
2.6 |
0.2 |
0.6 |
6.9 |
|
Frizzled |
|||||||
NM_021457 |
Fzd1 |
8.0 |
17.0 |
1.0 |
1.0 |
2.2 |
|
Induced by LPS in BMM | |||||||
|---|---|---|---|---|---|---|---|
Class A |
|||||||
NM_009912 |
Ccr1 |
0.9 |
3.1 |
7.4 |
0.8 |
0.7 |
|
NM_009911 |
Cxcr4 |
0.3 |
0.1 |
7.2 |
0.7 |
0.1 |
|
NM_004230 |
Edg5 |
0.2 |
0.5 |
2.0 |
0.4 |
0.6 |
|
NM_182806 |
Gpr18 |
17.8 |
52.2 |
10.8 |
1.0 |
1.6 |
|
NM_175493 |
Gpr68 |
3.3 |
1.7 |
40.2 |
1.5 |
0.8 |
|
NM_013533 |
Gpr162 |
1.1 |
0.5 |
6.7 |
0.8 |
0.8 |
|
NM_008311 |
Htr2b |
0.6 |
0.5 |
15.8 |
0.6 |
0.3 |
|
NM_008519 |
Ltb4r1 |
0.8 |
0.8 |
7.5 |
0.9 |
0.7 |
|
NM_008773 |
P2ry2 |
3.7 |
7.5 |
3.7 |
1.9 |
1.1 |
|
NM_021381 |
Prokr1 |
0.8 |
37.4 |
7.2 |
0.3 |
0.1 |
|
NM_008964 |
Ptger2 |
2.4 |
1.3 |
11.7 |
0.3 |
0.1 |
|
NM_008967 |
Ptgir |
1.1 |
2.3 |
7.4 |
0.7 |
0.6 |
|
Class B |
|||||||
NM_139138 |
Emr4 |
1.1 |
2.0 |
1.0 |
1.0 |
0.9 |
|
NM_001081298 |
Lphn2 |
2.2 |
1.4 |
1.2 |
0.3 |
0.3 |
|
Induced by LPS in TEPM | |||||||
|---|---|---|---|---|---|---|---|
Class A |
|||||||
NM_007577 |
C5r1 |
1.1 |
0.7 |
0.1 |
3.0 |
1.1 |
|
NM_007722 |
Cxcr7 |
0.5 |
0.5 |
0.5 |
1.0 |
4.0 |
|
NM_133200 |
P2ry14 |
0.7 |
1.7 |
1.4 |
0.8 |
6.7 |
|
GPCRs repressed by LPS in macrophages.
Relative Normalised Expression |
|||||||
|---|---|---|---|---|---|---|---|
Receptor Class |
Accession Number |
Gene Name |
BMM |
TEPM |
|||
2 h |
6 h |
24 h |
1 h |
7 h |
|||
Repressed by LPS in BMM and TEPM | |||||||
Class A |
|||||||
NM_009924 |
Cnr2 |
0.2 |
0.1 |
0.3 |
0.3 |
0.1 |
|
NM_009911 |
Cxcr4 |
0.3 |
0.1 |
7.1 |
0.7 |
0.1 |
|
NM_183031 |
Ebi2 |
0.2 |
0.1 |
0.1 |
0.4 |
0.2 |
|
NM_004230 |
Edg5 |
0.2 |
0.5 |
2.0 |
0.4 |
0.6 |
|
NM_022320 |
Gpr35 |
0.2 |
0.4 |
1.6 |
0.6 |
0.1 |
|
NM_030258 |
Gpr146 |
0.1 |
0.4 |
1.0 |
0.4 |
0.1 |
|
NM_008772 |
P2ry1 |
0.3 |
0.1 |
0.2 |
0.3 |
0.1 |
|
NM_175116 |
P2ry5 |
0.1 |
0.1 |
0.7 |
0.3 |
0.1 |
|
NM_013641 |
Ptger1 |
0.7 |
0.5 |
0.3 |
0.7 |
0.3 |
|
NM_008965 |
Ptger4 |
3.1 |
0.5 |
1.0 |
1.9 |
0.3 |
|
Repressed by LPS in BMM | |||||||
|---|---|---|---|---|---|---|---|
Class A |
|||||||
NM_007420 |
Adrb2 |
0.3 |
0.1 |
0.1 |
1.9 |
0.8 |
|
NM_009779 |
C3ar1 |
0.7 |
0.5 |
0.3 |
0.8 |
0.6 |
|
NM_007577 |
C5r1 |
1.1 |
0.7 |
0.1 |
3.0 |
1.1 |
|
NM_009915 |
Ccr2 |
0.5 |
0.4 |
0.2 |
0.8 |
0.9 |
|
NM_009910 |
Cxcr3 |
0.7 |
0.1 |
0.1 |
1.6 |
0.9 |
|
NM_009987 |
Cx3cr1 |
0.5 |
0.1 |
0.1 |
1.0 |
0.7 |
|
NM_030720 |
Gpr84 |
8.3 |
10.0 |
0.2 |
7.7 |
50.0 |
|
NM_027571 |
P2ry12 |
0.5 |
0.1 |
0.2 |
1.0 |
1.0 |
|
Class B |
|||||||
NM_018782 |
Calcrl |
1.0 |
2.6 |
0.2 |
0.6 |
7.1 |
|
Repressed by LPS in TEPM | |||||||
|---|---|---|---|---|---|---|---|
Class A |
|||||||
NM_007719 |
Ccr7 |
1.0 |
5.0 |
1.9 |
0.4 |
2.9 |
|
NM_021381 |
Prokr1 |
0.8 |
33.3 |
7.1 |
0.3 |
0.1 |
|
NM_198168 |
Ppp2r5b |
1.0 |
1.3 |
1.1 |
0.8 |
0.5 |
|
NM_008964 |
Ptger2 |
2.4 |
1.3 |
11.1 |
0.3 |
0.1 |
|
Class B |
|||||||
NM_080437 |
Celsr3 |
0.8 |
1.2 |
2.3 |
0.2 |
0.2 |
|
NM_173036 |
Gpr97 |
0.8 |
1.0 |
0.8 |
0.5 |
0.2 |
|
NM_016894 |
Ramp1 |
1.0 |
1.0 |
1.0 |
0.5 |
0.2 |
|
Class C |
|||||||
NM_022420 |
Gprc5b |
1.0 |
1.0 |
1.3 |
0.8 |
0.1 |
|
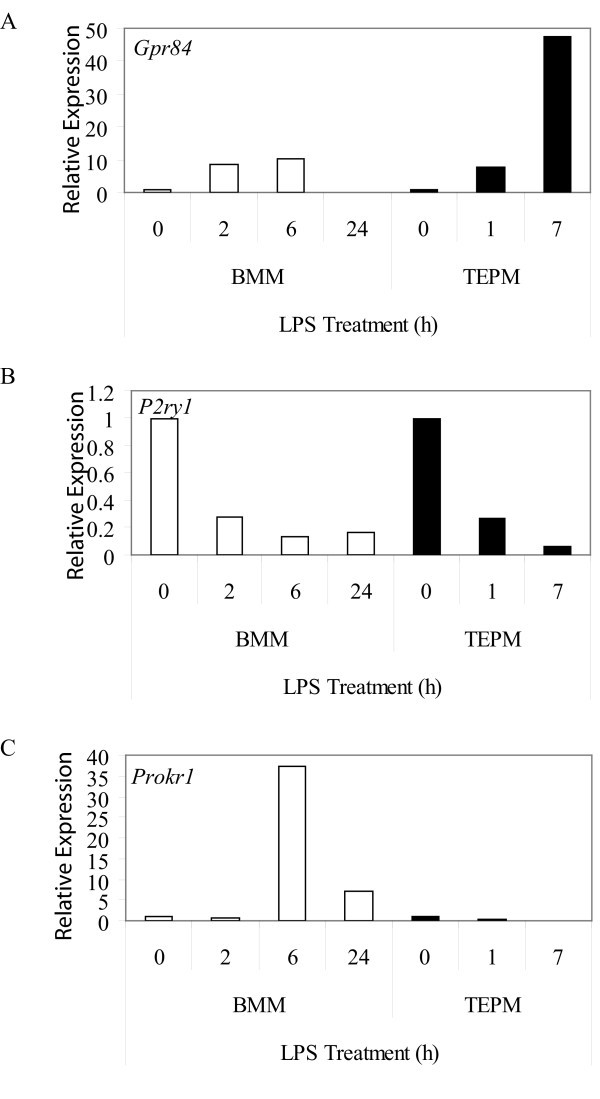
GPCRs regulated by LPS in macrophages. Micro-array expression analysis of Gpr84 (A), P2ry1 (B) and Prokr1 (C) mRNA in BMM and TEPM over a timecourse of 0, 2, 6, and 24 h, and 0, 1 and 7 h, respectively. Data points show gene expression relative to untreated control for each cell population (0 h).
The LPS time courses studied in BMM (2, 6 and 24 h) and TEPM (1 and 7 h) were not identical and so it is difficult to make definitive conclusions about non-conserved regulation by LPS in these two cell populations. Nonetheless, we did identify genes that appeared to be differentially regulated by LPS in BMM or TEPM at similar time points. Gpr18, P2ry2, P2ry12 and Adrb2 were all strongly regulated by LPS in BMM, but not TEPM (Table 3, 4), whilst LPS regulated Gprc5b expression in TEPM, but not BMM (Table 4). The effects of LPS could even be opposing in these two cell populations; Prokr1 was induced 37-fold by LPS in BMM (6 h), but repressed 20-fold in TEPM (7 h) (Figure 3c). Again some of these differences may be explained by the proliferative state of the cells; LPS triggers growth arrest in BMM and so some of the LPS-regulated genes unique to BMM may reflect this phenomenon. This is clearly not the case for all genes however (e.g. P2ry14 and Prokr1), thus arguing that subtle differences in LPS signaling pathways may exist in BMM versus TEPM.
Discussion
With an understanding of the GPCRs that are (a) highly expressed by the mononuclear phagocyte system, (b) selectively expressed by certain macrophage populations, or (c) regulated by the pro-inflammatory stimulus LPS, one can make some inferences about the role of specific GPCRs in macrophage biology. One of the most obvious findings from our analysis was that many of the P2Y purinergic receptors, which detect purine and pyrimidine nucleotides, were expressed in a restricted and/or regulated fashion in macrophages. Mechanical stress, cellular injury, inflammatory stimuli such as LPS, degranulation of mast cells and hypoxia can all increase extracellular concentrations of ATP and UTP [17, 18, 19, 20]. Consequently, these receptors are very likely to impact upon macrophage-mediated inflammatory responses.
Of the P2Y family, perhaps most striking in its expression pattern was P2ry6, which was essentially restricted to cells of the myeloid lineage. P2ry6 mRNA levels were greatly elevated in BMM, TEPM, RAW264 cells, microglia, osteoclasts and dendritic cells, compared to all other cell types (Figure 1b). P2ry6 expression in osteoclasts, which are a closely related lineage to macrophages, has been confirmed by others [21]. Whilst no previous studies have indicated that P2ry6 expression is restricted to the monocyte/macrophage lineage, there is evidence to suggest that this receptor does regulate the function of macrophages. UDP, which is reportedly a selective P2RY6 agonist [22, 23], triggered interleukin (IL)-8 release from the human monocyte-like cell line, THP-1 [24]. Conversely, the P2RY6 receptor antagonists, anthraquinone-sulfonic acid derivative reactive blue 2 and suramin, inhibited LPS-induced IL-8 production by THP-1 cells [24]. In microglia, P2ry6 mRNA expression was up-regulated in response to neuronal damage and UDP promoted phagocytosis by microglia, thus implying a role for P2RY6 in the clearance of damaged or dead neuronal cells [23]. Our observation that this receptor is widely expressed in the mononuclear phagocyte system suggests that this receptor may have a broader function in sensing and engulfing damaged cells throughout the body, as well as promoting pro-inflammatory responses in response to LPS and/or UDP.
This laboratory previously reported that P2ry5 mRNA expression was dramatically repressed by LPS in human and mouse macrophages [2], a finding supported by micro-array data presented here. This receptor was originally classified as a member of the P2Y receptor family on the basis of sequence identity and its ability to bind the nucleotides ATP and ADP [25]. Nucleotides failed to initiate downstream signaling from this receptor however [26], and it is therefore possible that P2RY5 acts as an antagonist of other P2Y receptors and restricts the macrophage response to extracellular nucleotides. Down-regulation of P2RY5 in response to a macrophage activating stimulus such as LPS might remove this inhibitory effect, thus allowing activated macrophages to respond to nucleotides. Alternatively, it is possible that the initial assignment of P2RY5 as a P2 receptor was incorrect. Cross-genome phylogenetic analysis showed that it clustered with a heterogenous group of receptors including the protease-activated receptors, as well as lysophosphatidylcholine (LPA) and sphingosylphosphorylcholine (S1P) receptors, rather than with other P2Y receptors [27]. In this case, LPS-mediated macrophage activation would presumably render cells unresponsive to the true ligand for this receptor.
Emerging evidence also implicates other P2Y receptors in macrophage-mediated inflammatory responses. For example, extracellular ATP, an agonist of P2RY2, induced IL-6 transcription in human monocyte-derived macrophages [28] and chemotaxis in human neutrophils [29], while the P2RY1 agonist 2-methylthio ATP increased IL-6 secretion from mouse splenocytes [30]. In support of a role for P2Y ATP receptors in macrophage function, and consistent with our data, P2ry1 and P2ry2 mRNAs were expressed at high levels in macrophages and LPS-stimulated monocytes [31]. Apart from these P2Y family members, P2RY12 was also reported to be selectively expressed in microglia in the rat brain [32]. Our analysis also showed that this receptor was highly enriched in microglia, as well as BMM, plasmacytoid dendritic cells, osteoclasts and a variety of lymphoid and neural tissues [see additional file 2a]. Finally, the P2Y-like receptor, P2ry14 (Gpr105), which is a receptor for UDP-conjugated sugars, mediated chemotaxis of bone marrow hematopoietic stem cells [33]. Despite the emerging literature that suggests a role for P2Y receptors in macrophage-mediated migration and inflammatory responses, a clear understanding of the selective agonists of these receptors and their interplay with each other and non-P2Y receptors is lacking. Nonetheless, our demonstration of the restricted and regulated expression of this family in macrophages identifies them as promising candidates for further analysis.
Other GPCRs that had a relatively restricted expression pattern and were strongly expressed by macrophages include Edg5, Gpr85 and Gpr84 (Table 2, Figure 1a and 1c, Additional file 2b). Edg5 is one of eight members of the endothelial differentiation gene (EDG) family of GPCRs that recognize LPA and S1P [34]. Several studies have documented potent effects of the EDG5 ligand, S1P on macrophages. Rat alveolar macrophages responded to S1P by producing O2 - at levels comparable to those induced by LPS or formyl-Methionyl-Leucyl-Phenylalanine [35]. Not surprisingly, given this finding, anti-microbial effects of S1P have been reported. In human monocyte-derived macrophages or THP-1 cells infected with M. smegmatis or M. tuberculosis, S1P reduced intracellular bacterial loads in a dose-dependent manner. Similar effects of S1P were also apparent in mice infected with M. smegmatis or M. tuberculosis [36]. Apart from activating anti-microbial responses, S1P also provided a survival signal to both mouse and human macrophage cell lines upon subsequent exposure to an apoptosis-inducing signal [37]. EDG1 was implicated in this response, but given the high expression of Edg5 reported here, this receptor may also be involved in macrophage responses to S1P.
Expression of the SREB (super conserved receptor expressed in brain) family member Gpr85 (Sreb2) was restricted to macrophages and neuronal tissues (Figure 1a). Whilst the expression of Gpr85 in the central nervous system was previously reported [38, 39, 40], a function has yet to be identified. The co-regulated expression of receptors such as Gpr85 in neural tissues and leukocyte populations may provide a mechanism for the neuro-immune cross-talk that has been observed in several studies [41, 42].
Gpr84 is an orphan GPCR that was strikingly restricted to BMM and microglia in an unstimulated state (Table 2, Figure 1c) and was strongly up-regulated by LPS in both BMM and TEPM (Figure 3a, Table 3). This up-regulation is consistent with elevated Gpr84 expression in microglia and tissue macrophages in an endotoxin shock model [43]. Microglia also expressed high levels of Gpr84 in the experimental autoimmune encephalomyelitis model of multiple sclerosis [43]. These data imply that GPR84 has a role in neuroinflammation, but the exact function is difficult to predict without any knowledge of a ligand. Nonetheless, given the highly restricted expression pattern of Gpr84 mRNA, antibodies against this receptor may provide useful tools for tracking macrophage populations in vivo.
As with Gpr84, mRNA for several other GPCRs, particularly chemokine receptors, were differentially expressed between various unstimulated macrophage populations. TEPM expressed elevated levels of mRNA for Cxcr4, Ccr1, Ccrl2, whilst BMM expressed higher levels of Ccr2, Cx3cr1 and Cxcr3. The fractalkine receptor CX3CR1 has been identified as a marker of patrolling monocytes that survey the endothelial surface [44] and its down-regulation on macrophages that have already trafficked to an inflammatory site is not surprising. Conversely, CXCR4 has a well-characterized function in targeting leukocytes and their progenitors away from the circulation [45], consistent with its enhanced expression in TEPM. Apart from chemokine receptors, the expression of a number of other GPCRs was different between BMM and TEPM. For example, the thrombin-related receptor, Ebi2 [46], was strongly expressed in BMM but not TEPM, whilst Gprc5b was enriched in TEPM but not BMM. Such differences are likely to contribute to functional divergence between tissue macrophage and inflammatory macrophage populations.
Many of the GPCRs that were highly expressed by macrophages in an unstimulated state (described above), were also regulated by LPS. Several other GPCRs were weakly expressed by unstimulated macrophages but were strongly regulated by LPS. One such example was the frizzled family member, Fzd1. Frizzleds (Fzd) represent the cell membrane receptors for a family of secreted glycoproteins called wingless-related proteins (WNTs). Wnts play essential roles in development, including cell fate determination, adhesion, polarity, migration and proliferation [47, 48, 49, 50]. The WNT family has also been implicated in immune regulation. For example, the WNT family member, WNTD, acted as a negative feedback regulator of Toll signaling in Drosophila [51]. In human macrophages, WNT5A expression was up-regulated by TLR ligands, as well as challenge with M. tuberculosis and M. avium, and the WNT5A/Frizzled-5 (FZD5) pathway promoted IL-12 production from peripheral blood mononuclear cells [52]. The striking up-regulation of Fzd1 in BMM (17-fold at 6 h LPS) implies that this FZD family member shares similar functions in regulating macrophage inflammatory responses.
We also identified several GPCRs that were differentially regulated by LPS in BMM and TEPM. In BMM, Prokr1 was transiently induced by LPS (37-fold at 6 h) but repressed in TEPM (20-fold at 7 h). PROKR1 is a receptor for prokineticin 1 (PK1), a peptide that regulates monocyte differentiation, as well as macrophage activation and migration. PK1 triggered the differentiation of both murine and human bone marrow progenitor cells into the monocyte/macrophage lineage [53], and reprogrammed the response of human monocytes to LPS by amplifying IL-12 and tumor necrosis factor alpha (TNF) production, but inhibiting IL-10 release [53]. The differential regulation of Prokr1 by LPS in BMM versus TEPM implies that this receptor has distinct functions during activation of tissue versus inflammatory macrophage populations.
Gpr18 was induced 52-fold in BMM following 6 h LPS exposure, while it remained unchanged in TEPM. This receptor was also highly expressed in other immune cell populations including B cells, T cells and DC cells [see additional file 2c]. N-arachidonylglycine (NAGly), a conjugate of arachidonic acid and glycine, is an endogenous ligand for GPR18 [54] that was reported to suppress inflammatory pain and have analgesic properties [55, 56]. Given the potent regulation of Gpr18 expression in BMM by LPS, NAGly is also likely to regulate macrophage function. The adenosine receptor, Adora2b was also strongly induced by LPS in BMM (191-fold at 6 h) yet was only modestly regulated (3-fold at 7 h) in TEPM. Adenosine-mediated activation of this receptor augmented IL-10 production by RAW264 cells in response to LPS [57]. In contrast, adenosine attenuated LPS-induced production of the pro-inflammatory cytokines TNF [58, 59, 60, 61] and IL-12 [60, 62] in mouse and human macrophages. Inducible Adora2b expression may therefore contribute to feedback regulation of macrophage inflammatory responses.
Conclusion
In this study, we have documented several GPCRs that are detectably expressed in primary mouse macrophages (BMM, TEPM), enriched in either or both BMM and TEPM, or regulated in activated macrophages (LPS-stimulated BMM and/or TEPM). For some of these genes, constitutive or regulated expression in macrophages has not previously been described. Future studies on such GPCRs and their agonists are likely to provide important insights into macrophage biology, as well as novel inflammatory pathways which could be future targets for drug discovery.
Methods
Cell culture and reagents
All cell lines and tissues were sourced from 8–10 week old male C57Bl/6 mice, with the exception of female-specific organs, which were sourced from female mice. All procedures were carried out in accordance with local guidelines for animal research. For female tissues, material was pooled from three females and for each female an average four embryos resulting in four umbilical cords and placentas were obtained. For other tissues, material was derived from a pool of three males. Biological replicates were defined as independent RNA preparations from independent pools of mice. Technical replicates were defined as independent amplifications from the same RNA sample. The full list of cell lines and tissues profiled, as well as further details on biological versus technical replication, is provided in additional file 3 (key to figures 1, 2 and additional file 2).
Mouse BMM were prepared from bone marrow cells from femurs of adult C57Bl/6 mice by culturing for 7 days in complete medium (RPMI containing 10% FCS, 20 U/ml penicillin, 20 μg/ml streptomycin and 2 mM L-glutamine (Invitrogen) in the presence of 10 000 U/ml CSF-1 (a gift from Chiron, Emeryville, CA) on bacteriological plastic plates. On day 7, BMM were treated with 10 ng/mL Salmonella minnesota LPS (Re595 mutant, Sigma) for 2, 6 or 24 h, and RNA was extracted. To generate TEPM, cells were harvested from the peritoneal cavity of C57Bl/6 mice 3 d after i.p. administration of 3% thioglycollate broth (Difco). Cells were then plated onto tissue culture plastic for 2 h, after which time non-adherent cells were removed by washing with PBS. Adherent TEPM were then treated with LPS for 1 or 7 h, or were left untreated, and RNA was extracted. Microglia were generated from cultures of mixed cortical cells. After 10 d culture, microglia were separated from astroglia by shaking on an orbital shaker (180 rpm, 20 min, 37°C) in the presence of 12 mM lidocaine (Sigma).
RNA and cDNA preparation
RNA extraction from mammalian cells or tissues was performed using RNeasy kits (Qiagen) or the standard Trizol protocol. If tissue amounts were more than 50 mg per mouse when dissected, tissues were pulverized while frozen. RNA was prepared separately for every mouse in order to identify samples with potentially degraded RNA. Trizol-extracted RNA was purified with a Qiagen RNeasy column. For BMM, osteoblasts and osteoclasts contaminating genomic DNA was removed during the RNeasy cleanup using DNaseI (Qiagen). The integrity and concentration of RNA was determined via microfluidic analysis on a bio-analyser (Agilent Technologies) or by analysis on a BioRad Experion (BioRad). Pooling occurred at the RNA level. For samples containing more than two μg total RNA available after pooling, standard Affymetrix single amplification was performed using two μg total RNA. For pooled samples containing less than two μg total RNA, 100 ng total RNA (or 50 ng for thymocyte SP CD8+) was used in a standard Affymetrix double amplification protocol. The starting quantity of RNA for specific tissues is included in additional file 3.
Microarray analysis
Standard Affymetrix protocols were used to process Affymetrix MOE430_2 microarrays. All CEL file images were processed as a single group using gcRMA [63, 64], after which the data for each gene was rescaled using per-gene normalisation to the median (for tissue specificity analysis) or per-gene normalisation to untreated control samples (for analysis of BMM and TEPM gene regulation by LPS). The microarray data described have been deposited in NCBIs Gene Expression Omnibus [5] and are accessible through GEO Series accession number GSE10246. GPCRs identified as highly enriched in BMM or TEPM were defined as transcripts where the normalised expression value was as least 10-fold greater in BMM/TEPM when compared to the median expression across all cell types and where BMM/TEPM expression significantly deviated from this median value (p-value < 0.05, student's two-tailed t-test). GPCRs regulated by LPS in primary mouse macrophages were defined as those whose normalised expression values were two-fold or greater up- or down-regulated at any point in the time course (2, 6 or 24 h LPS in BMM and/or 1 or 7 h LPS in TEPM) compared to the unstimulated state (0 LPS). LPS regulated genes were also filtered on the basis of statistical significance (p-value < 0.05, student's two-tailed t-test). All statistical testing was performed using the GeneSpring GX software, with the cross-gene error model function enabled.
Abbreviations
- BMM:
-
bone marrow-derived macrophage
- EDG:
-
endothelial differentiation gene
- Fzd:
-
Frizzled
- GPCR:
-
G Protein-Coupled Receptor
- IL:
-
interleukin
- LPA:
-
lysophosphatidylcholine
- LPS:
-
lipopolysaccharide
- NAGly:
-
N-arachidonylglycine
- P2Y:
-
purinergic receptor family Y
- PK1:
-
prokineticin 1
- S1P:
-
sphingosylphosphorylcholine
- SREB:
-
super conserved receptor expressed in brain
- TEPM:
-
thioglycollate-elicited peritoneal macrophage
- TLR:
-
Toll-like Receptor
- TNF:
-
tumor necrosis factor alpha
- Wnt:
-
Wingless-related protein.
Declarations
Acknowledgements
This work was supported by NHMRC Dora Lush and Queensland Government Smart State Fellowships to JL. A portion of this work was supported by the Novartis Research Foundation.
Authors’ Affiliations
References
- McKnight AJ, Macfarlane AJ, Dri P, Turley L, Willis AC, Gordon S: Molecular cloning of F4/80, a murine macrophage-restricted cell surface glycoprotein with homology to the G-protein-linked transmembrane 7 hormone receptor family. J Biol Chem 1996,271(1):486–489.View ArticlePubMedGoogle Scholar
- Lattin J, Zidar DA, Schroder K, Kellie S, Hume DA, Sweet MJ: G-protein-coupled receptor expression, function, and signaling in macrophages. J Leukoc Biol 2007,82(1):16–32.View ArticlePubMedGoogle Scholar
- Su AI, Wiltshire T, Batalov S, Lapp H, Ching KA, Block D, Zhang J, Soden R, Hayakawa M, Kreiman G, Cooke MP, Walker JR, Hogenesch JB: A gene atlas of the mouse and human protein-encoding transcriptomes. Proc Natl Acad Sci U S A 2004,101(16):6062–6067.View ArticlePubMedGoogle Scholar
- Genomics Institute of the Novartis Research Foundation SA: Genomics Institute of the Novartis Research Foundation, [http://symatlas.gnf.org/SymAtlas]. Google Scholar
- NCBIs Gene Expression Omnibus : NCBIs Gene Expression Omnibus, [http://www.ncbi.nlm.nih.gov/geo/]. Google Scholar
- Foord SM, Bonner TI, Neubig RR, Rosser EM, Pin JP, Davenport AP, Spedding M, Harmar AJ: International Union of Pharmacology. XLVI. G protein-coupled receptor list. Pharmacol Rev 2005,57(2):279–288.View ArticlePubMedGoogle Scholar
- International Union of Basic and Clinical Pharmacology GPCRRFF: International Union of Basic and Clinical Pharmacology, [http://www.iuphar-db.org/GPCR/ReceptorFamiliesForward]. Google Scholar
- Austyn JM, Gordon S: F4/80, a monoclonal antibody directed specifically against the mouse macrophage. Eur J Immunol 1981,11(10):805–815.View ArticlePubMedGoogle Scholar
- Hirsch S, Austyn JM, Gordon S: Expression of the macrophage-specific antigen F4/80 during differentiation of mouse bone marrow cells in culture. J Exp Med 1981,154(3):713–725.View ArticlePubMedGoogle Scholar
- Chenoweth DE, Goodman MG, Weigle WO: Demonstration of a specific receptor for human C5a anaphylatoxin on murine macrophages. J Exp Med 1982,156(1):68–78.View ArticlePubMedGoogle Scholar
- Gasque P, Singhrao SK, Neal JW, Wang P, Sayah S, Fontaine M, Morgan BP: The receptor for complement anaphylatoxin C3a is expressed by myeloid cells and nonmyeloid cells in inflamed human central nervous system: analysis in multiple sclerosis and bacterial meningitis. J Immunol 1998,160(7):3543–3554.PubMedGoogle Scholar
- Shimada T, Matsumoto M, Tatsumi Y, Kanamaru A, Akira S: A novel lipopolysaccharide inducible C-C chemokine receptor related gene in murine macrophages. FEBS Lett 1998,425(3):490–494.View ArticlePubMedGoogle Scholar
- Katsuyama M, Ikegami R, Karahashi H, Amano F, Sugimoto Y, Ichikawa A: Characterization of the LPS-stimulated expression of EP2 and EP4 prostaglandin E receptors in mouse macrophage-like cell line, J774.1. Biochem Biophys Res Commun 1998,251(3):727–731.View ArticlePubMedGoogle Scholar
- Mandal P, Novotny M, Hamilton TA: Lipopolysaccharide induces formyl peptide receptor 1 gene expression in macrophages and neutrophils via transcriptional and posttranscriptional mechanisms. J Immunol 2005,175(9):6085–6091.PubMedGoogle Scholar
- Schaub A, Futterer A, Pfeffer K: PUMA-G, an IFN-gamma-inducible gene in macrophages is a novel member of the seven transmembrane spanning receptor superfamily. Eur J Immunol 2001,31(12):3714–3725.View ArticlePubMedGoogle Scholar
- Wahl JR, Goetsch NJ, Young HJ, Van Maanen RJ, Johnson JD, Pea AS, Brittingham A: Murine macrophages produce endothelin-1 after microbial stimulation. Exp Biol Med (Maywood) 2005,230(9):652–658.Google Scholar
- Bergfeld GR, Forrester T: Release of ATP from human erythrocytes in response to a brief period of hypoxia and hypercapnia. Cardiovasc Res 1992,26(1):40–47.View ArticlePubMedGoogle Scholar
- Bodin P, Burnstock G: Synergistic effect of acute hypoxia on flow-induced release of ATP from cultured endothelial cells. Experientia 1995,51(3):256–259.View ArticlePubMedGoogle Scholar
- Bodin P, Burnstock G: Increased release of ATP from endothelial cells during acute inflammation. Inflamm Res 1998,47(8):351–354.View ArticlePubMedGoogle Scholar
- Detwiler TC, Feinman RD: Kinetics of the thrombin-induced release of adenosine triphosphate by platelets. Comparison with release of calcium. Biochemistry 1973,12(13):2462–2468.View ArticlePubMedGoogle Scholar
- Korcok J, Raimundo LN, Du X, Sims SM, Dixon SJ: P2Y6 nucleotide receptors activate NF-kappaB and increase survival of osteoclasts. J Biol Chem 2005,280(17):16909–16915.View ArticlePubMedGoogle Scholar
- Communi D, Parmentier M, Boeynaems JM: Cloning, functional expression and tissue distribution of the human P2Y6 receptor. Biochem Biophys Res Commun 1996,222(2):303–308.View ArticlePubMedGoogle Scholar
- Koizumi S, Shigemoto-Mogami Y, Nasu-Tada K, Shinozaki Y, Ohsawa K, Tsuda M, Joshi BV, Jacobson KA, Kohsaka S, Inoue K: UDP acting at P2Y6 receptors is a mediator of microglial phagocytosis. Nature 2007,446(7139):1091–1095.View ArticlePubMedGoogle Scholar
- Warny M, Aboudola S, Robson SC, Sevigny J, Communi D, Soltoff SP, Kelly CP: P2Y(6) nucleotide receptor mediates monocyte interleukin-8 production in response to UDP or lipopolysaccharide. J Biol Chem 2001,276(28):26051–26056.View ArticlePubMedGoogle Scholar
- Webb TE, Kaplan MG, Barnard EA: Identification of 6H1 as a P2Y purinoceptor: P2Y5. Biochem Biophys Res Commun 1996,219(1):105–110.View ArticlePubMedGoogle Scholar
- Li Q, Schachter JB, Harden TK, Nicholas RA: The 6H1 orphan receptor, claimed to be the p2y5 receptor, does not mediate nucleotide-promoted second messenger responses. Biochem Biophys Res Commun 1997,236(2):455–460.View ArticlePubMedGoogle Scholar
- Metpally RP, Sowdhamini R: Cross genome phylogenetic analysis of human and Drosophila G protein-coupled receptors: application to functional annotation of orphan receptors. BMC Genomics 2005, 6:106.View ArticlePubMedGoogle Scholar
- Hanley PJ, Musset B, Renigunta V, Limberg SH, Dalpke AH, Sus R, Heeg KM, Preisig-Muller R, Daut J: Extracellular ATP induces oscillations of intracellular Ca2+ and membrane potential and promotes transcription of IL-6 in macrophages. Proc Natl Acad Sci U S A 2004,101(25):9479–9484.View ArticlePubMedGoogle Scholar
- Chen Y, Corriden R, Inoue Y, Yip L, Hashiguchi N, Zinkernagel A, Nizet V, Insel PA, Junger WG: ATP release guides neutrophil chemotaxis via P2Y2 and A3 receptors. Science 2006,314(5806):1792–1795.View ArticlePubMedGoogle Scholar
- Straub RH, Pongratz G, Gunzler C, Michna A, Baier S, Kees F, Falk W, Scholmerich J: Immunoregulation of IL-6 secretion by endogenous and exogenous adenosine and by exogenous purinergic agonists in splenic tissue slices. J Neuroimmunol 2002,125(1–2):73–81.View ArticlePubMedGoogle Scholar
- Guerra AN, Fisette PL, Pfeiffer ZA, Quinchia-Rios BH, Prabhu U, Aga M, Denlinger LC, Guadarrama AG, Abozeid S, Sommer JA, Proctor RA, Bertics PJ: Purinergic receptor regulation of LPS-induced signaling and pathophysiology. J Endotoxin Res 2003,9(4):256–263.PubMedGoogle Scholar
- Sasaki Y, Hoshi M, Akazawa C, Nakamura Y, Tsuzuki H, Inoue K, Kohsaka S: Selective expression of Gi/o-coupled ATP receptor P2Y12 in microglia in rat brain. Glia 2003,44(3):242–250.View ArticlePubMedGoogle Scholar
- Lee BC, Cheng T, Adams GB, Attar EC, Miura N, Lee SB, Saito Y, Olszak I, Dombkowski D, Olson DP, Hancock J, Choi PS, Haber DA, Luster AD, Scadden DT: P2Y-like receptor, GPR105 (P2Y14), identifies and mediates chemotaxis of bone-marrow hematopoietic stem cells. Genes Dev 2003,17(13):1592–1604.View ArticlePubMedGoogle Scholar
- Takuwa Y, Takuwa N, Sugimoto N: The Edg family G protein-coupled receptors for lysophospholipids: their signaling properties and biological activities. J Biochem (Tokyo) 2002,131(6):767–771.Google Scholar
- Hornuss C, Hammermann R, Fuhrmann M, Juergens UR, Racke K: Human and rat alveolar macrophages express multiple EDG receptors. Eur J Pharmacol 2001,429(1–3):303–308.View ArticlePubMedGoogle Scholar
- Garg SK, Volpe E, Palmieri G, Mattei M, Galati D, Martino A, Piccioni MS, Valente E, Bonanno E, De Vito P, Baldini PM, Spagnoli LG, Colizzi V, Fraziano M: Sphingosine 1-phosphate induces antimicrobial activity both in vitro and in vivo. J Infect Dis 2004,189(11):2129–2138.View ArticlePubMedGoogle Scholar
- Weigert A, Johann AM, von Knethen A, Schmidt H, Geisslinger G, Brune B: Apoptotic cells promote macrophage survival by releasing the antiapoptotic mediator sphingosine-1-phosphate. Blood 2006,108(5):1635–1642.View ArticlePubMedGoogle Scholar
- Hellebrand S, Schaller HC, Wittenberger T: The brain-specific G-protein coupled receptor GPR85 with identical protein sequence in man and mouse maps to human chromosome 7q31. Biochim Biophys Acta 2000,1493(1–2):269–272.PubMedGoogle Scholar
- Hellebrand S, Wittenberger T, Schaller HC, Hermans-Borgmeyer I: Gpr85, a novel member of the G-protein coupled receptor family, prominently expressed in the developing mouse cerebral cortex. Brain Res Gene Expr Patterns 2001,1(1):13–16.View ArticlePubMedGoogle Scholar
- Matsumoto M, Beltaifa S, Weickert CS, Herman MM, Hyde TM, Saunders RC, Lipska BK, Weinberger DR, Kleinman JE: A conserved mRNA expression profile of SREB2 (GPR85) in adult human, monkey, and rat forebrain. Brain Res Mol Brain Res 2005,138(1):58–69.View ArticlePubMedGoogle Scholar
- Salzet M: Invertebrate molecular neuroimmune processes. Brain Res Brain Res Rev 2000,34(1–2):69–79.View ArticlePubMedGoogle Scholar
- Ransohoff RM, Liu L, Cardona AE: Chemokines and chemokine receptors: multipurpose players in neuroinflammation. Int Rev Neurobiol 2007, 82:187–204.View ArticlePubMedGoogle Scholar
- Bouchard C, Page J, Bedard A, Tremblay P, Vallieres L: G protein-coupled receptor 84, a microglia-associated protein expressed in neuroinflammatory conditions. Glia 2007,55(8):790–800.View ArticlePubMedGoogle Scholar
- Auffray C, Fogg D, Garfa M, Elain G, Join-Lambert O, Kayal S, Sarnacki S, Cumano A, Lauvau G, Geissmann F: Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science 2007,317(5838):666–670.View ArticlePubMedGoogle Scholar
- Kucia M, Jankowski K, Reca R, Wysoczynski M, Bandura L, Allendorf DJ, Zhang J, Ratajczak J, Ratajczak MZ: CXCR4-SDF-1 signalling, locomotion, chemotaxis and adhesion. J Mol Histol 2004,35(3):233–245.View ArticlePubMedGoogle Scholar
- Birkenbach M, Josefsen K, Yalamanchili R, Lenoir G, Kieff E: Epstein-Barr virus-induced genes: first lymphocyte-specific G protein-coupled peptide receptors. J Virol 1993,67(4):2209–2220.PubMedGoogle Scholar
- Clevers H: Wnt/beta-catenin signaling in development and disease. Cell 2006,127(3):469–480.View ArticlePubMedGoogle Scholar
- Cohen SM, Di Nardo S: Wingless: from embryo to adult. Trends Genet 1993,9(6):189–192.View ArticlePubMedGoogle Scholar
- Dierick H, Bejsovec A: Cellular mechanisms of wingless/Wnt signal transduction. Curr Top Dev Biol 1999, 43:153–190.View ArticlePubMedGoogle Scholar
- Klingensmith J, Nusse R: Signaling by wingless in Drosophila. Dev Biol 1994,166(2):396–414.View ArticlePubMedGoogle Scholar
- Gordon MD, Dionne MS, Schneider DS, Nusse R: WntD is a feedback inhibitor of Dorsal/NF-kappaB in Drosophila development and immunity. Nature 2005,437(7059):746–749.View ArticlePubMedGoogle Scholar
- Blumenthal A, Ehlers S, Lauber J, Buer J, Lange C, Goldmann T, Heine H, Brandt E, Reiling N: The Wingless homolog WNT5A and its receptor Frizzled-5 regulate inflammatory responses of human mononuclear cells induced by microbial stimulation. Blood 2006,108(3):965–973.View ArticlePubMedGoogle Scholar
- Dorsch M, Qiu Y, Soler D, Frank N, Duong T, Goodearl A, O'Neil S, Lora J, Fraser CC: PK1/EG-VEGF induces monocyte differentiation and activation. J Leukoc Biol 2005,78(2):426–434.View ArticlePubMedGoogle Scholar
- Kohno M, Hasegawa H, Inoue A, Muraoka M, Miyazaki T, Oka K, Yasukawa M: Identification of N-arachidonylglycine as the endogenous ligand for orphan G-protein-coupled receptor GPR18. Biochem Biophys Res Commun 2006,347(3):827–832.View ArticlePubMedGoogle Scholar
- Burstein SH, Rossetti RG, Yagen B, Zurier RB: Oxidative metabolism of anandamide. Prostaglandins Other Lipid Mediat 2000,61(1–2):29–41.View ArticlePubMedGoogle Scholar
- Huang SM, Bisogno T, Petros TJ, Chang SY, Zavitsanos PA, Zipkin RE, Sivakumar R, Coop A, Maeda DY, De Petrocellis L, Burstein S, Di Marzo V, Walker JM: Identification of a new class of molecules, the arachidonyl amino acids, and characterization of one member that inhibits pain. J Biol Chem 2001,276(46):42639–42644.View ArticlePubMedGoogle Scholar
- Nemeth ZH, Lutz CS, Csoka B, Deitch EA, Leibovich SJ, Gause WC, Tone M, Pacher P, Vizi ES, Hasko G: Adenosine augments IL-10 production by macrophages through an A2B receptor-mediated posttranscriptional mechanism. J Immunol 2005,175(12):8260–8270.PubMedGoogle Scholar
- Bouma MG, Stad RK, van den Wildenberg FA, Buurman WA: Differential regulatory effects of adenosine on cytokine release by activated human monocytes. J Immunol 1994,153(9):4159–4168.PubMedGoogle Scholar
- Hasko G, Szabo C, Nemeth ZH, Kvetan V, Pastores SM, Vizi ES: Adenosine receptor agonists differentially regulate IL-10, TNF-alpha, and nitric oxide production in RAW 264.7 macrophages and in endotoxemic mice. J Immunol 1996,157(10):4634–4640.PubMedGoogle Scholar
- Link AA, Kino T, Worth JA, McGuire JL, Crane ML, Chrousos GP, Wilder RL, Elenkov IJ: Ligand-activation of the adenosine A2a receptors inhibits IL-12 production by human monocytes. J Immunol 2000,164(1):436–442.PubMedGoogle Scholar
- McWhinney CD, Dudley MW, Bowlin TL, Peet NP, Schook L, Bradshaw M, De M, Borcherding DR, Edwards CK 3rd: Activation of adenosine A3 receptors on macrophages inhibits tumor necrosis factor-alpha. Eur J Pharmacol 1996,310(2–3):209–216.View ArticlePubMedGoogle Scholar
- Hasko G, Kuhel DG, Chen JF, Schwarzschild MA, Deitch EA, Mabley JG, Marton A, Szabo C: Adenosine inhibits IL-12 and TNF-[alpha] production via adenosine A2a receptor-dependent and independent mechanisms. Faseb J 2000,14(13):2065–2074.View ArticlePubMedGoogle Scholar
- Wu Z, Irizarry RA: Stochastic models inspired by hybridization theory for short oligonucleotide arrays. J Comput Biol 2005,12(6):882–893.View ArticlePubMedGoogle Scholar
- Zhijin Wu RAI Robert Gentleman,, Francisco Martinez-Murillo FS: A Model Based Background Adjustment forOligonucleotide Expression Arrays. Johns Hopkins University, Dept of Biostatistics Working Papers 2004.Google Scholar
- Flower DR: Modelling G-protein-coupled receptors for drug design. Biochim Biophys Acta 1999,1422(3):207–234.PubMedGoogle Scholar
- Gether U, Kobilka BK: G protein-coupled receptors. II. Mechanism of agonist activation. J Biol Chem 1998,273(29):17979–17982.View ArticlePubMedGoogle Scholar
- Pierce KL, Premont RT, Lefkowitz RJ: Seven-transmembrane receptors. Nat Rev Mol Cell Biol 2002,3(9):639–650.View ArticlePubMedGoogle Scholar
Copyright
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.